
Wysiłek fizyczny to podniesienie kosztu fizjologicznego w odpowiedzi na bodziec wysiłkowy (5). Jest to praca mięśni szkieletowych wraz z całym zespołem towarzyszących jej czynnościowych zmian w organizmie. Miarą bodźca jest jego: objętość, intensywność, cykliczność, częstotliwość (10).
Można mówić o:- wysiłku ogólnym – występuje wówczas, gdy w wysiłku zaangażowanych jest ponad 30% masy mięśniowej (zależy od masy ciała);
- wysiłku lokalnym – występuje wtedy, gdy w wysiłku zaangażowane jest mniej niż 30% całej masy ciała (praca 1 lub 2 kończyn);
- wysiłku maksymalnym– gdy zapotrzebowanie na tlen podczas wykonywania pracy jest równe indywidualnej wartości maksymalnej objętości pobranego podczas wysiłku tlen (5).
W czasie wysiłku fizycznego następuje przetwarzanie energii chemicznej powstającej w czasie metabolizmu komórkowego w energię mechaniczną niezbędną w pracy mięśni. Człowiek w czasie pracy fizycznej wykorzystuje do 30% energii zawartej w produktach żywnościowych. Pozostała część jest zamieniana w ciepło. Procesy metaboliczne związane są z przemianą materii i energii ,mogą przebiegać w warunkach tlenowych i beztlenowych (10). W wykonywaniu wysiłku kluczową rolę odgrywa praca mięśni. Mięśnie szkieletowe stanowią około 42 procent masy ciała człowieka (2) .
W spoczynku • 60% zapotrzebowania energetycznego mięśni pokrywane przez wolne kwasy tłuszczowe,
• 40%- przez glukozę
Podczas wysiłku max w ciągu niewielu milisekund zapotrzebowanie energetyczne włókien mięśniowych wzrasta ok. 100 –krotnie (10).
1. Klasyfikacja wysiłków (10):1. Ze względu na dominujący rodzaj skurczu• Wysiłek dynamiczny - mięśnie zmieniają długość a względnie stałe napięcie
(skurcz izotoniczny)
• Wysiłek statyczny - mięśnie zmieniają napięcie i nie zmienia się długość (skurcz izometryczny)
2. Ze względu na wielkość grup mięśni biorących udział w wykonywaniu pracy:• wysiłek lokalny- udział w wysiłku mniej niż 30% masy mięśni(np praca jedną kończyną)
• wysiłek ogólny- w wysiłku bierze udział ponad 30% masy mięśniowej
3. Ze względu na rodzaj toru przemian energetycznych:• Tlenowe – aerobowe
• Beztlenowe – anaerobowe
1. fosfagenowe ( niekwasomlekowe);
2. glikolityczne ( kwasomlekowe)
• Mieszane- tlenowo-beztlenowe
4. W zależności od czasu wykonywania pracy:• wysiłek krótkotrwały - (do 15 min.)
• wysiłek o średnim czasie - czasie trwania (do 30 min.)
• wysiłek długotrwały - (ponad 30 min.)
Intensywność – obciążenia dzielimy z kolei na:• wysiłki maksymalne - zapotrzebowanie na tlen jest równe indywidualnej wartości (= 100% VO2 max).
• wysiłki submaksymalne zapotrzebowanie na tlen jest niższe niż (< 100%VO2 max).
do 20% VO2max - lekkie
20% - 50% VO2max - średnie
50 – 75% VO2max - ciężkie
> 75% VO2max - b. ciężkie
• wysiłki supramaksymalne zapotrzebowanie na tlen przekracza (>100% VO2 max
2. Energetyka wysiłku fizycznego (10,5).W wysiłku praca mięśni w 40% powoduje wytworzenie energii - ATP, oraz 60% w postaci ciepła. W wydajność energetyczna pracy mięśniowej (współczynnik pracy użytecznej) to proporcja między wielkością wykonanej pracy mechanicznej a ilością wydatkowanej energii chemicznej
Współczynnik pracy użytecznej wynosi przykładowo:Chód – ok. 35% (przy prędkości 4,5km/h )
Bieg - 25-30%
Jazda na rowerze - 20-28%
Praca małych grup mięśni - 10-15%
Energetyka kataboliczna różnych substratów: Proces utleniania:
1g kwasów tłuszczowych → 37,7 kJ
1g białek lub cukrów → 16,7 kJ
Ilość energii wytworzonej przy zużyciu 1 litra tlenu zależy od rodzaju substratu energetycznego kw. tłuszczowe węglowodany , a do określenia substratu energetycznego w wysiłku służy:
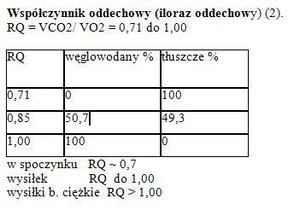
Parametry krążeniowe w spoczynku (7):1.Vw – objętość wyrzutowa serca (SV – volume stroke) to ilość krwi wtłaczana do zbiornika tętniczego w czasie 1 cyklu pracy serca.
Norma 60 – 100ml
Vw = 101 + 0,5Cs – 1,09Cr – 0,61 m
Vw – objętość wyrzutowa ( ml )
Cs –ciśnienie skurczowe
Cr – ciśnienie rozkurczowe
m – wiek w latach
2. V min –Q- pojemność minutowa serca (CO – cardiac output)- to ilość krwi tłoczona przez komorę serca w czasie 1 minuty
Norma 90ml/s = 5,4 l/min
Vmin = HR x Vw
Vmin - objętość minutowa serca ( ml/min)
HR – tętno ( uderzenia/min)
Vw – objętość wyrzutowa serca
3. HR – tętno – (heart rate) - faliste odkształcenie tętnicy podczas skurczu serca. Skurcz komór serca powoduje powstanie tzw. fali tętna w tętnicach.
Częstość tętna - ilość uderzeń serca na minutę.
U dorosłych ilość uderzeń na minutę wynosi ok. 60-80
U dzieci: 90-140/minutę
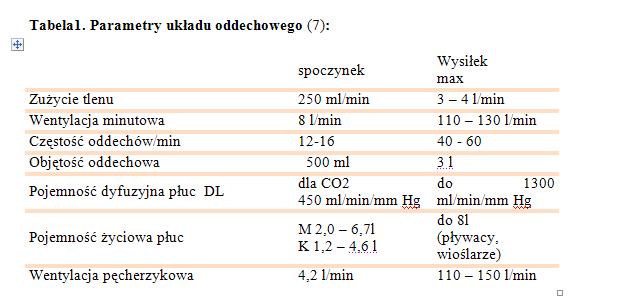
3.
Adaptacja – odgrywa niezwykle ważną rolę w wysiłku fizycznym, zwłaszcza w ekstremalnych warunkach. Określamy ją jako zdolność organizmu do przystosowania się do nowych warunków pod wpływem stresora. Polega na przystosowanie i przestrojenie funkcji do zapewnienia utrzymania stanu równowagi czynnościowej ( homeostazy) w nowej sytuacji(10).
Adaptacja kształtowana jest przez wysiłek i restytucję.
Bodziec wywołujący adaptację musi mieć odpowiednią
- intensywność (60 – 80% wywołujący adapt.) (30-60% podtrzymujący)
- czas trwania, częstotliwość
Restytucja to ustalenie nowej homeostazy organizmu po zakończeniu wysiłku fizycznego(10):Prawa restytucji:I. Heterochronizmu:
Genetycznie uwarunkowana kolejność wypoczywania poszczególnych układów: nerwowego, oddechowego, krążenia i odbudowy energetycznej.
II. Okresowości:
faza szybka do 5 min. po zakończeniu wysiłku - likwidacja długu tlenowego
faza wolna – w zależności od wysiłku trwa od kilku min do kilkunastu godzin cd restytucji:
III. Fazowości:
występują w czasie restytucji: faza kompensacji i może wystąpić faza superkompensacji
praca
Adaptacja do wysiłków długich(10):1. ↑ poj życiowej płuc
2. ↑ VO2max
3. ↑ poj. tlenowej krwi, liczba RBC, Hb, Ht, Mb w mięśniach
4. Przerost lewej komory serca ↑SV i Q – bradykardia spoczynkowa
5. W mięśniach : ↑ ukrwienia ( kapilaryzacji mięśni), ↑ włókien ST i ich objętości,↑liczby mitochondriów i grzebieni mitochondrialnych,↑ enzymów metabolizmu tlenowego
4. Koszt fizjologiczny wysiłku fizycznego (10,7,1).4.1.Koszt fizjologiczny różni się od pojęcia koszt energetyczny. Koszt energetyczny wykonywanej pracy możemy precyzyjnie zmierzyć. Obciążenie fizjologiczne związane jest z funkcjonowaniem narządów i układów człowieka związanych z charakterem wykonywanej pracy i nie ma obiektywnych metod, które by pozwalały je dokładnie określić. Ma ono cechy wysoce zindywidualizowane chociaż można wyodrębnić pewne wspólne uwarunkowania wynikające z charakteru wykonywanej pracy. Dla przykładu funkcje układu pokarmowego, krążenia i innych ma związek z charakterem wykonywanej pracy w sensie obciążenie jak i towarzyszącym stresem i emocjami. Trudno jest znaleźć bezpośredni związek miedzy tymi zależnościami chociaż związek taki niewątpliwie istnieje. Wydatek energetyczny jest często stosowany jako energetyczne kryteria ciężkości pracy fizycznej. Do oceny ciężkości pracy wykorzystywana jest wielkość wydatku energetycznego w ciągu zmiany roboczej.
4.2. Metody pomiaru wydatku energetycznego:-kalorymetria bezpośrednia
-kalorymetria pośrednia
-chronometrażowa (tabelaryczna wg Lehmana)
-oceny na podstawie mechanicznego efektu pracy
-oceny na podstawie zmian fizjologicznych
-oceny uciążliwości wynikającej z wysiłków statycznych
-oceny uciążliwości związanej z monotypowością ruchów roboczych
-badanie „odnowy tętna”
4.3. Metody tabelaryczne: przy braku możliwości wykonywania pomiaru wydatku energetycznego jedną z wymienionych wcześniej metod wartości te możemy określić za pomocą szacunkowej metody chronometrażowo-tabelarycznej, odczytując z tabeli wartość wydatku energetycznego dla typowych czynności.
4.4 Źródła substratów: zmiany zachodzące w mięśniach podczas krótkotrwałego wysiłku o dużej intensywności są taki, że wysiłek trwa zbyt krótko aby koszt energetyczny mógł być pokryty przez tlenowe procesy metaboliczne (spalanie węglowodanów i tłuszczy), dlatego podczas takiego wysiłku głównym źródłem energii są beztlenowe procesy metaboliczne. Są wówczas wykorzystywane mięśniowe zapasy ATP, fosfokreatyny i glikogenu. Zapasy te mogą się zmniejszać bardzo znacznie co obniża zdolność kompleksów aktomiozynowych do skracania się. Glukoza uwolniona z glikogenu jest metabolizowana do kwasu pirogronowego, który w warunkach braku tlenu zostaje zredukowany do kwasu mlekowego. Stężenie kwasu mlekowego w mięśniach gwałtownie rośnie, to z kolei powoduje wzrost stężenia jonów wodorowych w komórkach mięśniowych-my odczuwamy to jako zakwasy.
5. Praca mięśni a wysiłek fizyczny (1, 10):Skurcz mięśnia to podstawowe fizjologiczne wydarzenie, które umożliwia nam podejmowanie wysiłku fizycznego.
W organizmie człowieka wyróżnia się trzy rodzaje mięśni:
1. szkieletowe (łączą się ze szkieletem)
2. gładkie (tworzą mięśniową wyściółkę dla organów)
3. mięsień sercowy
W wysiłku fizycznym mowa jest o pracy mięśni szkieletowych, dlatego też w artykule tym skupimy się na nich.
5.1. Budowa mięśni szkieletowych (5,1): Każdy mięsień szkieletowy zbudowany jest z wielu cylindrycznych komórek mięśniowych zwanych włóknami (miofibryle).Każda miofibryla składa się z sarkomerów, natomiast sarkomer składa się z cieniutkich pręcików zwanych miofilamentami. Miofilamenty składają się z dwóch części. Grubych filamentów zbudowanych z miozyny oraz cienkich filamentów zbudowanych z aktyny. Kiedy biegamy grube i cienkie filamenty oddziałują wzajemnie między sobą wytwarzając skurcz mięśnia.Innym ważnym składnikiem komórki mięśniowej są mitochondria. Znajdują się one obok filamentów, a ich podstawową funkcją jest produkcja energii potrzebnej przy skurczu mięśniowym.W komórce mięśniowej znajdują się również kropelki tłuszczowe oraz glikogen.
5.2.Rodzaje włókien mięśniowychWyróżniamy dwa główne typy włókien mięśniowych:
-typ I wolnokurczliwe (ST)
-typ II szybkokurczliwe (FT)
Ich skróty pochodzą od angielskich słów "fast twich" co znaczy że mają dużą prędkość skracania i "slow twich" co znaczy że mają małą prędkość skracania.
Wśród włókien typu II można wyróżnić kilka podtypów: IIa (czerwone), IIb (białe), IIc (forma pośrednia między IIb i IIb).
Różnice w prędkości skracania zarówno jednych jak i drugich włókien są głównie wynikiem występowania w tych włóknach różnych form ATP-azy miozynowej. ATP-aza to enzym który powoduje rozpad ATP. Włókna ST mają formę wolną ATP-azy a włókna FT formę szybką. W organizmie człowieka występują zarówno włókna wolno jak i szybkokurczliwe a ich zawartość procentowa determinuje to czy jesteśmy bardziej wytrzymali, szybcy czy silni. Przewaga włókien wolnokurczliwych informuje o tym, że jesteśmy bardziej wytrzymali. Natomiast przewaga włókien szybkokurczliwych daje nam większe możliwości szybkościowo-siłowe.Warte uwagi jest również to, iż poszczególne mięśnie człowieka mają różne składy włókien.
5.3. Mechanizm skurczu mięśniaDziałanie mięśni szkieletowych polega na wytwarzaniu siły, która może być rozwinięta dopiero po otrzymaniu sygnału z ośrodkowego układu nerwowego. Impuls ten rozprzestrzenia się dzięki włóknom nerwowym i dochodzi do jednostki motorycznej, a tym samym do wszystkich jej włókien mięśniowych.Do skurczu mięśnia niezbędna jest energia, a jej bezpośrednim źródłem jest adenozynotrifosoforan(ATP)
5.4. Rodzaje skurczów mięśniowychPojedynczy – jest to pojedyncze pobudzenie mięśnia.
Tężcowy – następuje w momencie kiedy mięsień zostanie pobudzony serią bodźców w której przerwa między bodźcami jest krótsza niż cały czas skurczu.
5.5. Funkcje mięśni:Każdy skoordynowany ruch w naszym ciele wymaga zastosowania siły mięśniowej. Wykonanie tych ruchów możliwe jest tylko dzięki pracy mięśni, które ze względu na funkcje można podzielić na następujące grupy:
agonistyczne - grupa mięśni, których skurcz powoduje określony ruch np. zginanie,
antagonistyczne - grupa mięśni przeciwstawiających się agonistom,
synergistyczne – asystują mięśniom agonistycznym, stabilizują staw.
Na przykład zginanie w stawie łokciowym wymaga skracania mięśnia ramiennego i dwugłowego ramienia (biceps) - mm agonistyczne i rozluźnienia mięśnia trójgłowego ramienia (triceps) - antagonista. Mięsień ramienno-promieniowy (synergista) współdziała z mięśniem dwugłowym ramienia i mięśniem ramiennym, czyli asystuje w zginaniu w stawie łokciowym.
5.6. Warunki pracy mięśnia (2,5,10):Koncentryczne – podstawowa funkcja mięśni czyli skracanie odnosi się właśnie do działania w warunkach koncentrycznych. Podczas działania koncentrycznego mięśni, występuje ruch w stawie i zalicza się on do ruchów dynamicznych.
Statyczne – mięśnie mogą również pracować bez poruszania dźwigniami kostnymi
i wywoływania ruchu w stawie. Kiedy to następuje mięśnie rozwijają siłę, ale ich długość pozostaje bez zmiany (czyli jest stała). Warunki statyczne występują wtedy, kiedy próbujemy podnieść przedmiot, którego ciężar jest większy od siły naszych mięśni.
Ekscentryczne – występują wtedy, gdy działająca siła zewnętrzna jest większa niż możliwa do rozwinięcia i wtedy mięsień ulega wydłużeniu. Ruch ten nazywamy ekscentrycznym. Ponieważ w stawie występuje ruch, jest to więc również ruch dynamiczny. Jego przykładem jest praca mięśnia dwugłowego ramienia w chwili prostowania w stawie łokciowym, gdy ciężar, który powinieneś unieść przekracza możliwości utrzymania go. W tym momencie nie upuszczasz ciężaru gwałtownie na ziemię tylko próbujesz przeciwdziałać temu i następuje rozciąganie mięśnia. Tego rodzaju warunki występują podczas schodzenia po schodach, zbiegu z góry lub zeskoku z wysokości (Jaskólski 2002).
6. Energia a wysiłek fizyczny (3,5).6.1. Do pracy mięśni potrzebna jest energia. Bardzo specyficznym i będącym zarazem jedynym źródłem tej energii jest ATP (adenozynotrifosforan). Możemy powiedzieć, że nasze mięśnie są jak silnik zamieniający energię chemiczną w energię kinetyczną (ruch). Wartą zaznaczenia rzeczą jest fakt, iż mięśnie nie tylko zużywają, ale także produkują energię.
6.2. Systemy produkcji energii(3,11):
Różnica pomiędzy tymi systemami wynika z tego, iż organizm pozyskuje energię do resyntezy ATP z różnych źródeł.
1. Anaerobowy niekwasomlekowy – w systemie tym organizm nie wykorzystuje tlenu i nie produkuje kwasu mlekowego. A do resyntezy ATP (czyli ponownego odtworzenia tego związku) używa fosfokreatyny. Ten system uruchamia się przy wysiłkach maksymalnych trwających według różnych źródeł około 10 sekund. do produkcji „paliwa” pochodzącego z cukrów, tłuszczy i niewielkiej ilości białek potrzebuje tlenu. Produkcja kwasu mlekowego jest znikoma.
2. Anaerobowy kwasomlekowy - system ten uruchamia się przy wysiłkach trwających do 2min. Rozpad cząsteczek cukru zachodzi bez udziału tlenu ale organizm produkuje kwas mlekowy. Głównym substratem energetycznym są cukry.
7. Kwas mlekowy (4,10, 5).Wzrastające zakwaszenie w czasie wysiłków fizycznych tradycyjnie zwykło się tłumaczyć zwiększoną produkcją kwasu mlekowego, który uwalniając proton H+ doprowadza do spadku PH. Jeżeli ilość wolnego protonu H+ przekroczy wewnątrzkomórkowe systemy buforujące dochodzi do wzrostu zakwaszenia i objawowej kwasicy metabolicznej. Według tej teorii jedną z wielu przyczyn ograniczających możliwości wysiłkowe mięśnia w czasie intensywnej pracy jest właśnie wzrost stężenia mleczanu, prowadzący do wewnątrzkomórkowej kwasicy. Tradycyjna interpretacja bazuje na fakcie, iż kwas mlekowy jest stosunkowo silnym kwasem organicznym (pKa=3,87), który w środowisku komórkowym występuje w całkowicie zdysocjowanej formie. Uwolnione protony H+ doprowadzają właśnie do wystąpienia kwasicy metabolicznej zwanej również, od rzekomego dawcy protonów, kwasicą mleczanową.
Jako pierwszy istnienie kwasu mlekowego odkrył w 1780 roku szwedzki chemik Carl Wilhelm Scheele. Jego obecność stwierdził w próbkach kwaśnego mleka, co było powodem do nadania "nowej" cząsteczce nazwy kwas mlekowy (właściwa nazwa chemiczna to kwas 2-hydroksypropanowy). Dwaj laureaci nagrody Nobla w 1922 roku Otto Meyerhoff i Archibald V. Hill byli pionierami badań nad biochemią wysiłków fizycznych. Ich badania nad utlenianiem węglowodanów pokazały, iż w warunkach niedoboru tlenu utlenianie glukozy prowadzi do powstania kwasu mlekowego. Kwas mlekowy powstaje również w czasie kiedy nagle wzrasta zapotrzebowanie energetyczne pracującego mięśnia, a źródła dostarczania substratów energetycznych przy udziale procesów tlenowych nie są wystarczające. Stwierdzenie obecności kwasu mlekowego w pracującym mięśniu dało podstawę do wysnucia teorii o przyczynie rozwoju kwasicy metabolicznej, której to bezpośrednią przyczyną ma być narastające stężenie kwasu mlekowego. Na bazie tego poglądu przeprowadzano wiele eksperymentów, które miały potwierdzić założoną tezę. Wyniki jednego z nich zaprezentowano poniżej.
Przy dużej intensywności pracy, mięśnie produkują kwas mlekowy, który pojawia się we krwi w postaci soli nazywanej mleczanem. Kwas mlekowy i mleczan są powszechnie uznawane za istotną przyczynę wystąpienia zmęczenia, drętwienia i bolesności powysiłkowej wśród trenujących osób. Czy jest to jednak słuszne założenie? Autorzy ostatnich badań potwierdzili możliwość istnienia zmęczenia mięśni przy niskim poziomie mleczanu oraz niemalże braku zmęczenia przy jego wysokim stężeniu. Dowodzą również, iż w wielu sytuacjach możliwości wysiłkowe biegacza nie zależą, jak sądzi wielu, od produkcji mniejszej ilości mleczanu, lecz na możliwości do jego wręcz większego wytwarzania. Podważają również istnienie progu anaerobowego (beztlenowego) – takiej intensywności wysiłku, powyżej której większa część energii pozyskiwana jest na drodze metabolizmu beztlenowego (glikoliza) – wskazując na fakt, że obecna wiedza naukowa nie potwierdza jego istnienia. Badacze sugerują, że nawet, jeśli próg anaerobowy istnieje, nie ma tak dużego znaczenia (zastosowania) jakie niektórzy mu przypisują. Szeroko uznawane przekonanie, że kwas mlekowy jest źródłem wszystkich dolegliwości jest wciąż popierane przez naukowców, którzy nie śledzą nowości albo nie mają odwagi, by powiedzieć prawdę sportowcom, gdyż boją się przeciwstawić popularnej opinii. Jeśli zbadamy jednak szczegóły tego, jak energia jest produkowana w mięśniach w różnego rodzaju wysiłkach, stwierdzamy, iż mleczan nie jest odpowiedzialny za dolegliwości, za które niektórzy go obwiniają.
Mit 1: Skurcze mięśnia są powodowane obecnością kwasu mlekowego w mięśniu.Przykurcz nie jest spowodowany akumulacją kwasu mlekowego. Można bowiem zaobserwować występowanie przykurczy zarówno przy wysokiej koncentracji mleczanu, jak również kiedy stężenie mleczanu w mięśniach jest niskie. Nocą kiedy ludzie często cierpią z powodu skurczów mięśniowych, koncentracja mleczanu we krwi jest mała. Z drugiej strony w czasie biegu na dystansie 400m, gdzie wszyscy zawodnicy kończą bieg z mleczanem we krwi na poziomie 18-25 razy wyższym niż poziom spoczynkowy, przykurcze są rzadkością. W wielu przypadkach przykurcze występują podczas nasilonych wysiłków o długim czasie trwania. W takich warunkach koncentracja mleczanu jest prawdopodobnie wyraźnie wyższa niż w spoczynku, lecz dużo poniżej maksymalnych wartości obserwowanych podczas bardzo intensywnych, ale krótkich wysiłków. Dlatego nie można winić kwasu mlekowego, faktu jego nagromadzenia za występowanie przykurczy. Powodowane one są nadwrażliwością tkanki mięśniowej oraz nadreaktywnością układu nerwowego wysyłającego impulsy pobudzające.
Mit 2:Obecność kwasu mlekowego w mięśniu powoduje sztywność i bolesność mięśniowąOpóźniony początek mięśniowej bolesności (z ang. delayed onset of muscle soreness) to ból, który pojawia się dzień lub dwa dni po nietypowym, intensywnym wysiłku. Ten typ bólu pojawia się głównie po ćwiczeniach ze znaczną ilością skurczy ekscentrycznych. Są to skurcze mięśni przy jednoczesnym ich wydłużaniu się (np. amortyzacja upadku). Te bóle mięśniowe nie mają nic wspólnego z obecnością kwasu mlekowego w mięśniach. Badania dostarczają dowodów na potwierdzenie tego faktu. W jednym z badań uczestnik miał do przebiegnięcia dwa testy: 9 x 5 minut z prędkością 3,5 m/s z dwuminutową przerwą, pierwszy po płaskim terenie, a drugi na 10%-owym pochyleniu. Bieganie po płaskim, gdzie odnotowano większą koncentrację mleczanu, nie powodowało bolesności. Natomiast dzień po bieganiu na pochyleniu, stwierdzono niższą koncentrację mleczanu, a badany odczuwał poważny ból. Powyższe doświadczenie dobrze znane jest tym, którzy biegają na górzystych trasach. To właśnie zbieganie, które wymaga większej ilości skurczów ekscentrycznych jest główną przyczyną bolesności powysiłkowej, nie zaś bieg po płaskim terenie czy też podbiegi. To powoduje więcej szkód w mięśniach, ponieważ liczba włókien mięśniowych zaangażowanych do skurczu o specyficznym napięciu jest 4-8 razy większa przy skurczu ekscentrycznym niż przy koncentrycznym. Dlatego w wyniku pracy większej ilości włókien mięśniowych powstają większe napięcia, a to powoduje mikrourazy i następujące stany zapalne. Jest to logiczne wytłumaczenie tego, że kwas mlekowy nie ma nic wspólnego z bolesnością powysiłkową mięśni.
Najbardziej przekonujące są sytuacje występowania zmęczenia przy niskim stężeniu mleczanu jak również braku zmęczenia przy jego wysokiej koncentracji. Na przykład, pod koniec szczególnie wymagającej konkurencji, jaką jest bieg na 100 kilometrów, poziom zmęczenia jest niezwykle wysoki, ale koncentracja mleczanu we krwi jest niewiele wyższa niż spoczynkowa. U ludzi z opisanym syndromem McArdle’a stwierdzono znacznego stopnia ograniczenie możliwości wysiłkowych, a przecież praktycznie nie dochodzi do powstawania kwasu mlekowego w ich komórkach mięśniowych. Zatem zmęczeniu mięśniowemu może towarzyszyć bardzo niski poziom kwasu mlekowego albo wręcz jego całkowity brak. Obserwując poziom kwasicy w mięśniach, zauważamy jej znaczny wzrost podczas skurczów izometrycznych, co może potwierdzać hipotezę, iż kwas mlekowy jest odpowiedzialny za wywołanie zmęczenia. Wykonując taki wyczerpujący wysiłek izometryczny mięśnia czworogłowego uda (np. oparcie o ścianę plecami z nogami ugiętymi pod kątem prostym pomiędzy udem, a podudziem, tak jak byśmy siedzieli na krześle) doprowadzamy do sytuacji, kiedy postępujące zmęczenie spowoduje czasowy spadek generowanej mocy (osłabienie mięśnia). Po dwu minutowej przerwie obserwowane zmęczenie szybko maleje i niemal zupełnie zanika. Po tym czasie mięśnie mogą jeszcze raz generować początkową moc, pomimo tego, że podczas przerwy poziom mleczanu spada do wartości spoczynkowych raczej wolno utrzymując się nadal na wysokim poziomie. Stąd dwie minuty od zakończenia wysiłku ilość mleczanu w mięśniu jest jeszcze bardzo wysoka, pomimo tego mięśnie mogą jeszcze raz generować początkową moc, a samo zmęczenie w znacznym stopniu ustępuje. Dlatego obserwując sytuacje, kiedy występuje znaczna kwasica i duże stężenie mleczanu, a zmęczenie pozostaje na umiarkowanym poziomie, ciężko jest potwierdzić koncepcję, że wzrost poziomu kwasu mlekowego w mięśniach jest bezpośrednią przyczyną występowania czy nasilenia zmęczenia. Nic nie dowodzi temu, że mleczan jest bezpośrednim powodem, a już na pewno nie jest jedynym ani też głównym powodem występowania zmęczenia.
Niektórzy sądzą, że im więcej mleczanu produkujesz, tym jesteś mniej efektywny.
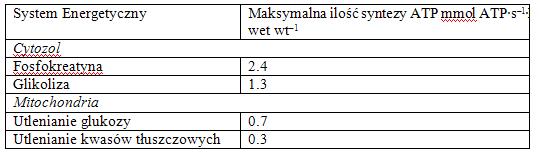
W rzeczywistości jest zupełnie odwrotnie. W wielu konkurencjach lekkoatletycznych, jeśli produkujesz więcej kwasu mlekowego, to znak, że pracujesz z większą intensywnością. Trzeba sobie również uświadomić, jakie są możliwości regeneracji zasobów energetycznych (ATP) komórki mięśniowej. Pomimo „dużej ekonomiki” przemian mitochondrialnych, ich możliwości dostarczania energii są znacznie ograniczone w stosunku do możliwości przemian glikolitycznych czy też reakcji kinazy kreatynowej. Dlatego możliwa jest sytuacja, kiedy nasilona praca doprowadza do wyczerpania możliwości dostarczania energii przez system mitochondrialny, a deficyt energetyczny pokrywany zostaje przez cytozolowe przemiany: glikolizę oraz reakcję kinazy kreatynowej.
 Kwasica metaboliczna
Kwasica metaboliczna powstaje w wyniku nasilonej utylizacji zasobów ATP połączonej ze wzrostem udziału glikolizy w resyntezie ATP. Na potrzeby pracy komórki mięśniowej zostaje uwolniona energia w czasie hydrolizy bogatoenergetycznego wiązania fosforanowego cząsteczki ATP. Powstałe ADP oraz reszta fosforanowa Pi są bezpośrednio włączane w cykl przemian glikolitycznych gdzie następuje resynteza ATP. W czasie glikolizy zredukowaniu ulega NAD+ do formy NADH oraz powstają protony. W warunkach wydolności mitochondrialnego systemu pozyskiwania energii powstały NADH oraz protony H+ są przekazywane na łańcuch oddechowy gdzie są utylizowane przy udziale tlenu. W czasie niedoboru tlenu lub znacznego zapotrzebowania na energię (bardzo nasilonej pracy mięśniowej), przekraczającego możliwości systemu mitochondrialnego, pula zredukowanego nukleotydu NADH jest reoksydowana w czasie reakcji przekształcenia pirogronianu w mleczan. W tej reakcji również są przyłączane dwa protony H+ co zapobiega lub opóźnia wystąpienie kwasicy.
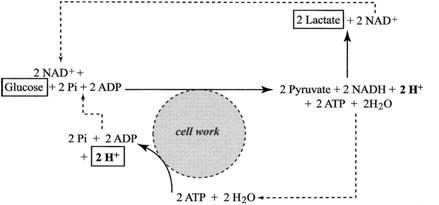
Jak widać z przedstawionego schematu (5) to niezbilansowane protony H+ powstające w wyniku hydrolizy ATP mają bezpośredni wpływ na kwasowość środowiska komórkowego. Można więc powiedzieć, iż kwasica metaboliczna rozwija się na skutek nasilenia przemian energetycznych komórki, a jej bezpośrednią przyczyną jest przebieg cytozolowych (pozamitochondrialnych) reakcji hydrolizy ATP.
W czasie hydrolizy ATP zostaje uwolniony proton H+ oraz reszta fosforanowa, która wykazuje duży potencjał do buforowania (przyłączania) protonów H+. Nie można jednak wyrównać (zbuforować) nadmiaru protonów H+ powstałych w reakcji hydrolizy ATP przez powstające jednoczenie reszty fosforanowe, gdyż te na bieżąco są zużywane do resyntezy ATP w czasie glikolizy oraz przez mitochondrialny łańcuch oddechowy. Ilość wolnych fosforanów jest więc nieproporcjonalnie mniejsza od ilości protonów H+ powstałych w wyniku hydrolizy ATP dlatego ich zdolność do buforowania jest znacznie mniejsza niż wynika to z przebiegu reakcji hydrolizy ATP.
Mówiąc o pozamitochondrialnych przyczynach kwasicy trzeba wiedzieć, że to właśnie ograniczenia przemian w obrębie mitochondriów doprowadzają do zaburzenia utylizacji protonów H+ i do następującego rozwoju kwasicy w wyniku ich nagromadzenia. W tak zwanym „stanie spoczynkowym”, kiedy zapotrzebowanie energetyczne komórki mięśniowej pozostaje na względnie niskim poziomie, nie dochodzi do kumulacji protonów gdyż te przekazywane są właśnie do mitochondriów gdzie są wykorzystywane do wytworzenia energii (ATP) w obrębie łańcucha oddechowego. Również pula zredukowanego nukleotydu NADH jest regenerowana w mitochondrium. Zostaje zachowana równowaga w bilansie dostarczania i utylizacji protonów H+. Wysilona praca mięśniowa niesie za sobą znaczny wzrost zapotrzebowania na energię. Dochodzi do przekroczenia możliwości na pokrycie potrzeb energetycznych przez system mitochondrialny, w wyniku czego znacznemu nasileniu poddawane są pozamitochondrialne szlaki energetyczne generujące ATP. Większa część ADP oraz fosforanów Pi jest włączana do glikolizy celem odtworzenia ATP. Wzrasta ilość utylizowanej glukozy, a nasilona glikoliza wyczerpuje dostępną pulę NAD+.
Reakcja dehydrogenazy mleczajowej (5) pozwala na odbudowanie zasobów NAD+ niezbędnych dla kontynuowania glikolizy w warunkach kiedy system mitochondrialny jest już niezdolny do przyjmowania „ponad miarę” substratów dla swoich przemian energetycznych.
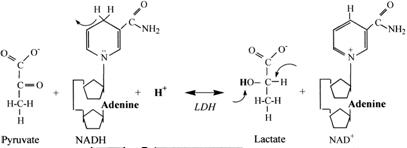
Odtworzona w tej reakcji pula NAD+ pozwala kontynuować glikolizę, a powstający mleczan opuszczając komórkę wraz z protonem H+ na drodze symportu stanowi swego rodzaju bufor czynnościowy dla gromadzących się protonów. Mleczan z krwi wyłapywany jest przez inne tkanki będące w stanie dodatniego bilansu energetycznego (głównie wątroba) gdzie włączany jest do przemian energetycznych: jako substrat dla glukoneogenezy (odtworzenie glukozy) lub jako paliwo po przekształceniu do pirogronianu i wprowadzeniu do cyklu Krebsa.
Kwasica metaboliczna jest zjawiskiem groźnym dla homeostazy organizmu bez względu na przyczynę jej powstawania. Z praktycznego punktu widzenia nie ma różnicy czy jest ona następstwem działania ATP-azy hydrolizującej ATP na potrzeby pracy mięśniowej, czy wynikiem nasilonej glikolizy. Kwasica jest niewątpliwie jednym z czynników ograniczających maksymalne wysiłki fizyczne. Ważne jednak aby prowadząc różnego rodzaju dywagacje na temat progu mleczanowego mieć świadomość, iż powstający mleczan sam w sobie nie jest zły. Nie jest tak, że organizmowi jest ciężko go „tolerować”, a to z kolei przyczynia się do rozwoju zmęczenia mięśniowego. Mleczan, jego droga powstawania, jest swego rodzaju wyjściem awaryjnym ratującym mechanizmy energetyczne przed całkowitym załamaniem. Warunkuje to możliwość przedłużenia okresu tak zwanej pracy na długu energetycznym oraz daje pełną niezależność od metabolizmu mitochondrialnego (tlenowego). W porównaniu do przemian cyklu Krebsa glikoliza jest znacznie bardziej wydajnym systemem dostarczania energii, zwłaszcza kiedy ta jest niezbędna szybko i w znacznych ilościach. Bilans mleczanu we krwi jest dobrym pośrednim wskaźnikiem zakwaszenia ustroju i daje swego rodzaju obraz o źródłach energetycznych zużywanych do pozyskania energii przy zadanej intensywności pracy. Kumulacja mleczanu świadczy bowiem o przekroczeniu możliwości dostarczania ATP przy pomocy metabolizmu mitochondrialnego, który jak wiadomo jest niezwykle ekonomiczny, a dla długodystansowca jest szczególnie ważną sprawą oszczędna gospodarka substratami węglowodanowymi ze względu na ich ograniczone możliwości magazynowania.
8. Wysiłek fizyczny a układ krążenia. Wpływ na ryzyko chorób sercowo-naczyniowych8.1. Wysiłek fizyczny należy do podstawowych elementów prewencji pierwotnej i wtórnej chorób układu krążenia, uwzględnionych w zaleceniach Europejskiego Towarzystwa Kardiologicznego. O możliwościach organizmu człowieka do wykonania określonego wysiłku fizycznego decydują mechanizmy adaptacyjne układu krążenia. Obejmują one zmiany: czynności serca i ciśnienia tętniczego krwi, objętości wyrzutowej i pojemności minutowej, różnicy tętniczo-żylnej wysycenia tlenem oraz dystrybucji przepływu krwi. Sprawność tych mechanizmów, rozstrzygająca o zdolności do wykonywania różnego rodzaju wysiłków fizycznych, ich efektywności oraz tolerancji zależą nie tylko od intensywności treningu ale także od określonych uwarunkowań genetycznych. Wśród nich wymienia się m.in. polimorfizm I/C genu ACE, polimorfizm R577X genu alfa aktyniny 3 ACTN3czy polimorfizm -9BDKBR12 genu baradykininy typu 2. Obecność określonych genotypów decyduje o większej zdolności do wykonywania albo intensywnego wysiłku siłowego, albo wytrzymałościowego. Jednocześnie na podstawie wielu badań, w tym jednego z pierwszych, opublikowanego w 1953 roku w piśmie Lancet, wiadomo, że regularny wysiłek fizyczny wpływa na zmniejszenie częstości zdarzeń sercowych, zgonów z powodu schorzeń układu krążenia i zgonów w ogóle. W świetle badań ostatnich lat okazuje się, że o oddziaływaniu wysiłku fizycznego na stan zdrowia człowieka również decydują uwarunkowania genetyczne. Wśród nich wymienia się m.in. polimorfizm V227A genu PPARalfa czy polimorfizm Arg16/Gly genu receptora adrenergicznego beta 2. Badania podłoża molekularnego wysiłku fizycznego mogą posłużyć do określenia genotypu, charakteryzującego ludzi, dla których wysiłek jest szczególnie ważnym i skutecznym sposobem zapobiegania chorobom układu krążenia. Każdy wysiłek fizyczny związany jest z określonymi zmianami w funkcjonowaniu układu krążenia, warunkującymi możliwość adaptacji do jego wykonania. Zmiany te są odmienne przy różnych wysiłkach. Rozróżniamy dwa podstawowe rodzaje wysiłku fizycznego: dynamiczny (izotoniczny), który wiąże się ze zmianą długości mięśni a nie ich napięcia oraz wysiłek statyczny (izometryczny) związany ze zmianą napięcia a nie długości mięśni. Oprócz rodzaju wysiłku, także jego intensywność, czas trwania czy warunki zewnętrzne, takie jak temperatura czy wilgotność powietrza, w których jest wykonywany, wpływają na przebieg adaptacji organizmu do zwiększonego obciążenia fizycznego. Procesy te zależą także w dużej mierze od sprawności różnych mechanizmów regulacyjnych oraz od uwarunkowań genetycznych. Te ostatnie czynniki wpływają nie tylko na możliwości adaptacyjne do aktualnie wykonywanego wysiłku, ale także decydują o możliwościach poprawienia sprawności i wydolności fizycznej w wyniku regularnego treningu. Są poza tym jednym z podstawowych elementów wpływających na skuteczność aktywności fizycznej jako elementu prewencji chorób sercowo-naczyniowych(7).
8.2. Zmiany czynnościowe zachodzące w układzie krążenia podczas wysiłku fizycznego:Adaptacja układu krążenia do wysiłku fizycznego obejmuje przyspieszenie czynności serca (HR), wzrost objętości wyrzutowej i pojemności minutowej, zwiększenie różnicy tętniczo-żylnej wysycenia krwi tlenem oraz ciśnienia tętniczego(12).Zmiany te mają na celu zaopatrzenie pracujących mięśni w odpowiednią ilość tlenu (7).
Adaptacja układu krążenia do wysiłku fizycznego obejmuje zmiany:– Czynności serca i ciśnienia tętniczego krwi
– Objętości wyrzutowej
– Pojemności minutowej
– Różnicy tętniczo-żylnej wysycenia tlenem
– Dystrybucji przepływu krwi
Czynność serca przyspiesza się wraz z trwaniem wysiłku fizycznego aż po kilku minutach dochodzi do ustalenia stanu równowagi ze stałą wartością HR, która zmienia się przy dalszym zwiększeniu intensywności wysiłku, by znów po 2-4 minutach ustabilizować się na określonym poziomie. Z reakcją tą, określaną w piśmiennictwie anglojęzycznym mianem steady state, mamy do czynienia podczas wysiłków dynamicznych submaksymalnych (chód, bieg, jazda na rowerze). Gdy wysiłek wykonywany jest w wysokich temperaturach wzrost częstości skurczów serca zwiększa się stale, co stanowi jeden z mechanizmów termoregulacyjnych. Oba mechanizmy mogą doprowadzić do osiągnięcia maksymalnej wartości HR, która określana jest w przybliżeniu wg prostego wzoru: 220 – wiek.
Zmiany ciśnienia tętniczego podczas wysiłku fizycznego dotyczą wyraźnego wzrostu ciśnienia skurczowego do wartości powyżej 200 mmHg proporcjonalnego do wzrostu intensywności wysiłku oraz w mniejszym stopniu wzrostu ciśnienia rozkurczowego – w warunkach prawidłowych max o 12% w stosunku do wartości w spoczynku(7).
Kolejnym parametrem ulegającym zwiększeniu w trakcie wysiłku jest objętość wyrzutowa. Na jej wartość ma wpływ współdziałanie pompy sercowej (warunkującej przepływ krwi przez pracujące mięśnie) i pompy obwodowej (utrzymanie powrotu żylnego, zapewniającego rozkurczowe wypełnienie serca), co ma szczególne znaczenie w wysiłku wykonywanym w pozycji wyprostnej. Objętość wyrzutowa wynosi u dorosłego człowieka średnio 80 ml w pozycji siedzącej lub stojącej i 110 ml w pozycji leżącej(7).
Na stopień wzrostu rozkurczowego wypełnienia komór serca mają wpływ(7):– wyjściowe napięcie włókien mięśnia serca;
– zwiększony dopływ krwi żylnej (skurcz pracujących mięśni szkieletowych);
– „efekt ssący” LK (różnica ciśnień między LK a LP we wczesnej fazie rozkurczu);
– redystrybucja krwi.
Kluczowe znaczenie w adaptacji układu krążenia do wysiłku odgrywa pompa obwodowa (9). Opisane czynniki decydujące o wartości objętości wyrzutowej stanowią też ograniczenie możliwości jej zwiększania, co oznacza, że wraz ze zwiększaniem intensywności wysiłku, początkowo dochodzi do stopniowego wzrostu objętości wyrzutowej, a następnie do jej ustalenia na stałym poziomie.
Wzrost objętości wyrzutowej i czynności serca decyduje o zwiększaniu wraz z trwaniem wysiłku fizycznego pojemności minutowej serca. Proces ten postępuje aż do osiągnięcia poziomu intensywności wysiłku odpowiadającej 40-60% maksymalnego pobierania tlenu (VO2max). Od tego momentu narastanie pojemności minutowej zależy już w przeważającym stopniu od przyspieszania czynności serca.
Różnica tętniczo-żylna wysycenia krwi tlenem wzrasta z średnio 5 ml O2/100 ml w spoczynku do 15 ml O2//100 ml w wysiłku maksymalnym, co jest efektem stopniowego obniżania się zawartości tlenu w krwi żylnej w wyniku zwiększenia ekstrakcji tlenu z krwi przepływającej przez pracujące mięśnie.
8.3. Kolejnym zjawiskiem, które wpływa na adaptację organizmu do wysiłku fizycznego jest
redystrybucja przepływu krwi przez różne obszary naczyniowe, w wyniku rozszerzenia naczyń krwionośnych w pracujących mięśniach szkieletowych i zwężenia łożyska naczyniowego w narządach wewnętrznych .Decydujące o tym mechanizmy to: aktywacja układu współczulnego oraz wzrost tempa lokalnego metabolizmu z gromadzeniem produktów przemiany materii (mleczanów, adenozyny, jonów wodorowych, jonów potasu, CO2), wzrostem temperatury mięśni, hipoksją i zwiększeniem ciśnienia osmotycznego. Podczas wysiłku fizycznego 80-85% pojemności minutowej trafia do mięśni szkieletowych, 4-5-krotnie wzrasta przepływ wieńcowy oraz o 30% wzrasta przepływ mózgowy(8).
O właściwej adaptacji układu krążenia do wysiłku fizycznego decydują mechanizmy regulacyjne, które można podzielić na wewnętrzne i zewnętrzne. Wewnętrzne to przede wszystkim układ bodźcoprzewodzący serca, zewnętrzne zaś to wszystkie wpływy nerwowe i hormonalne wynikające z aktywności układu współczulnego i przywspółczulnego (wpływ na HR, kurczliwość i szybkość przewodzenia) a także ośrodka naczynioruchowego rdzenia przedłużonego, który modyfikuje działanie układu autonomicznego w odpowiedzi na bodźce z receptorów obwodowych naczyń krwionośnych, mięśni i stawów (mechano-, baro-, proprio-receptory) oraz na impulsy z kory mózgowej (11).
8.4. Wpływ czynników genetycznych na zdolność do wykonywania wysiłków fizycznych.Badania ostatnich lat coraz wyraźniej wskazują, że zdolność do wykonywania wysiłku fizycznego, jego tolerancji oraz efektywność zależą nie tylko od intensywności treningu ale także od określonych uwarunkowań o charakterze genetycznym. Od nich także uzależnione są zmiany, jakie zachodzą w organizmie człowieka podczas wykonywania wysiłku fizycznego a także pod wpływem długotrwałego treningu. Czynniki genetyczne warunkują różną zdolność człowieka do wykonywania wysiłków siłowych bądź wytrzymałościowych. Wreszcie, jak się wydaje decydują również o skuteczności oddziaływania aktywności fizycznej jako czynnika zmniejszającego ryzyko chorób sercowo-naczyniowych(11).
Główny nurt badań, podobnie jak w innych dziedzinach medycyny, skierowany jest tutaj w stronę poszukiwania zależność między określoną cechą fizyczną a obecnością polimorfizmów genetycznych. Polimorfizm, czyli jednoczesne występowanie dwóch różnych kopii tego samego genu, powstałych w wyniku mutacji w jednej z nich, niejednokrotnie prowadzi do zmiany właściwości kodowanego przez ten gen białka. Polimorfizm może dotyczyć jednego nukleotydu (mutacja punktowa) lub dłuższych odcinków genu, ulegających delecji lub insercji. Zidentyfikowanie polimorfizmu określonego genu nie wystarcza do stwierdzenia jego związku z fenotypem – konieczne są badania statystyczne, wykazujące związek tego miejsca w genomie z występowaniem określonej cechy oraz badania funkcjonalne, potwierdzające zmianę właściwości genu i białka. Polimorfizm funkcjonalny może prowadzić do zmian ekspresji określonego genu, co możliwe jest do stwierdzenia np.: przy pomocy analizy poziomu mRNA metodą Nothern blot. Badania zmierzające do zidentyfikowania genów wpływających na tolerancję oraz efektywność wysiłku fizycznego w dużej mierze koncentrują się na grupie sportowców. Jak dotąd, mimo usilnych starań nie udało się określić tzw. „genów mistrzów”, nie mniej wiadomo już, że możliwość osiągania wybitnych wyników w sporcie jest najprawdopodobniej uwarunkowana genetyczne(8).
Szczególne zainteresowanie w tym zakresie wzbudza polimorfizm I/D (insercja/delecja 287 nukleotydowego fragmentu sekwencji niekodującej) genu enzymu konwertującego angiotensynę ACE. Wiadomo, że u osób z genotypem I/I stwierdza się niższy poziom krążącego oraz tkankowego stężenia ACE. Już wcześniej obserwowano, że wspomniany fenotyp jest częstszy wśród sportowców wyczynowych (11). Niemniej, bardzo ciekawych obserwacji dokonali w tym zakresie Thompson J i wsp. (8) wśród alpinistów. Przeprowadzone przez nich badania wskazują na istotną statystycznie różnicę genotypu genu ACE w obrębie polimorfizmu I/D na rzecz częstszego występowania genotypu I/I u alpinistów, którym udało się zdobyć szczyty powyżej 8000 m, w porównaniu do tych, którzy tego nie dokonali. Różnic tych nie obserwowano już między tymi spośród badanych alpinistów, którzy wchodzili powyżej 8000 m, bez dodatkowego źródła tlenu a tymi, którzy je stosowali. Z kolei w badaniach prowadzonych w Południowej Korei nie stwierdzano innej częstość występowania poszczególnych alleli tego genu wśród sportowców w porównaniu do grupy kontrolnej, choć wskazywano na wyraźnie większą częstość allelu I wśród biegaczy długodystansowych (9).
Badania polimorfizmu I/C genu ACE wśród sportowców, uprawiających różne dyscypliny, pokazały inną częstość występowania poszczególnych alleli u osób zdolnych do krótkotrwałych siłowych lub długotrwałych wytrzymałościowych wysiłków fizycznych (8). Na podstawie tych i innych badań wysunięto hipotezę, że allel D genu ACE odpowiada za zdolność do wykonywania krótkotrwałego wysiłku, wymagającego użycia większej siły, allel I natomiast predysponuje do wysiłków długotrwałych o charakterze wytrzymałościowym.
Wydaje się jednak, że zależność ta może być odmienna w różnych populacjach. W opublikowanych w 2007 roku badaniach polimorfizmu I/D genu ACE, przeprowadzanych wśród sportowców izraelskich, stwierdzono, że genotyp D/D występuje znacząco częściej u biegaczy długodystansowych niż wśród sprinterów (11). Zaprzecza to hipotezie na temat związku allelu I ze zdolnością do wysiłków wytrzymałościowych lub wskazuje, że ostateczny efekt polimorfizmu I/D genu ACE jest odmienny w różnych populacjach, prawdopodobnie w wyniku oddziaływania z innymi, nieznanymi zmianami na poziomie DNA.
Obecnie trudno więc jednoznacznie określić zależność zdolności do wykonywania wysiłku fizycznego od polimorfizmu I/C genu ACE. Wydaje się jednak, że ma on u człowieka istotne znaczenie dla przebiegu wysiłku fizycznego, prawdopodobnie w związku z odmiennym wpływem różnych stężeń enzymu konwertującego na metabolizm komórek mięśni szkieletowych oraz mięśnia serca.
Rola aktywności fizycznej w prewencji pierwotnej chorób układu krążenia jest obecnie niezaprzeczalna. Jedne z pierwszych badań wskazujących na korzystny wpływ ruchu na zdrowie to badanie opublikowane w 1953 roku w piśmie Lancet oceniające częstość epizodów niedokrwienia mięśnia serca oraz związanych z nimi zgonów w obserwacji 3-dniowej wśród pracowników londyńskiego transportu (11). Stwierdzono wyraźną różnicę w zakresie ocenianych zdarzeń między grupą konduktorów i kierowców, wiążąc ją z odmiennym trybem pracy. Siedząca praca kierowców była wiązana z większym narażeniem tej grupy zawodowej na zdarzenia o charakterze ostrego zespołu wieńcowego i związany z nimi zgon w pierwszych dniach obserwacji.
Z czasem pojawiło się wiele badań potwierdzających kluczowe znaczenie aktywności fizycznej w ograniczaniu ryzyka chorób sercowo-naczyniowych, w tym Harvard Alumni Health Study czy Interheart (9, 11).
Wnioski wynikające z powyższych obserwacji, chociaż bardzo dobitnie wskazywały na korzyści wynikające z codziennej aktywności fizycznej, w kolejnych latach zyskały bardzo ważne uzupełnienia. Dziś już wiemy, że bez porównania większe korzyści w prewencji chorób układu krążenia przynosi wysiłek fizyczny wykonywany w sytuacji odpoczynku i relaksu, w wolnym czasie, niż ten związany z wykonywaną pracą (8). Wydaje się, że znaczenie odgrywa tu psychiczne nastawienie i cel, w jakim podejmuje się aktywność fizyczną a nie jedynie sam fakt jej uprawiania. Jest to zgodne z aktualnym obecnie poglądem, że nie tyle sam ruch, co wynikające z niego dobre ogólne samopoczucie stanowi najważniejszy element pierwotnej prewencji chorób układu krążenia (11).
Warto zwrócić także uwagę na fakt, że szczególnymi beneficjentami zdrowego stylu życia są mężczyźni (ryc. 5). Na podstawie przeprowadzonych pod koniec lat 80-tych obserwacji stwierdzono, że mężczyźni nieaktywni fizycznie mają znacznie większe ryzyko zgonu z powodu chorób układu krążenia niż kobiety prowadząc podobny tryb życia (8). Dlatego właśnie dla nich zmiana stylu życia ma największe znaczenie.
W ostatnim czasie zwraca się również uwagę na korzyści w prewencji chorób układu krążenia wynikające z utrzymywania aktywności fizycznej przez osoby starsze. Poza tym są dowody wskazujące, że aktywność fizyczna zmniejsza ryzyko nie tylko chorób układu krążenia, ale także cukrzycy, nowotworów, depresji czy osteoporozy (11).
Istnieje wiele przekonywujących badań, wskazujących, że korzystne oddziaływanie wysiłku fizycznego na układ krążenia ma swoje uzasadnienie biologiczne. Stwierdzono m.in., że regularna aktywność fizyczna prowadzi do wielu zmian, takich jak zwiększenie stężenia cholesterolu HDL i zmniejszenie stężenia triglicerydów we krwi, obniżenie ciśnienia tętniczego, zmniejszenie insulinooporności i poprawę profilu glikemii. Na poziomie komórkowym dochodzi przede wszystkim do zwiększenia syntezy tlenku azotu oraz zmniejszenia stresu oksydacyjnego. Stwierdzono, że regularna aktywność fizyczna zapewnia utrzymanie równowagi między produkcją i inaktywacją tlenku azotu oraz przywraca zdolności regeneracyjnych krążących komórek progenitorowych. Trening fizyczny obniża również poziom białka C-reaktywnego, co może wyjaśniać jego korzystny wpływ na zmniejszenie ryzyka chorób układu krążenia i metabolicznych (8,9).
8.5. Wpływ uwarunkowań genetycznych na efekty biologiczne wysiłku fizycznego:Na podstawie prowadzonych w ostatnich latach badań można stwierdzić, że uwarunkowania genetyczne decydują również o wpływie wysiłku fizycznego na stan zdrowia.
Najbardziej podstawowych w tym zakresie informacji dostarcza badanie HERITAGE, w którym zbadano pod względem bardzo różnych parametrów zdrowotnych członków 205 rodzin (w sumie 793 osoby) po 20- -tygodniowym okresie ćwiczeń fizycznych. W badaniu tym stwierdzono między innymi, że poprawa profilu lipidowego, wtórna do zwiększenia aktywności fizycznej zależy od wrodzonych uwarunkowań, na co wskazywała wyraźna korelacja tych zmian ze stopniem pokrewieństwa badanych osób (11).
Obserwacje te zostały potwierdzone w opublikowanych w 2007 roku badaniach Naito i wsp. (8). Stwierdzono, że zmiana poziomu cholesterolu w surowicy krwi zależna od zwiększenia aktywności fizycznej jest inna u osób z różnym genotypem w zakresie polimorfizmu V227A genu PPARalfa. U osób z genotypem A227 nie obserwowano żadnego wpływu ćwiczeń fizycznych na poziom cholesterolu, podczas gdy u osób z genotypem PPARalfa-WT stwierdzano wyraźny wzrost wartości HDL w stopniu zależnym od intensywności wysiłku.
PodsumowanieWysiłek fizyczny jest obecnie uważany za jeden z najważniejszych elementów prewencji pierwotnej i wtórnej chorób sercowo-naczyniowych. Poza bezpośrednim wpływem aktywności fizycznej na procesy komórkowe coraz bardziej podkreśla się rolę związanego z nią dobrego samopoczucia psychicznego, które samo w sobie zmniejsza ryzyko zachorowania. W oddziaływaniu wysiłku fizycznego na organizm człowieka kluczową rolę odgrywa sprawność mechanizmów adaptacyjnych, takich jak przyspieszenie czynności serca, zwiększenie objętości wyrzutowej czy redystrybucja przepływu krwi a także określone uwarunkowania genetyczne. Badania zmierzające do zidentyfikowania ich mają na celu określenie genotypu, który będzie charakteryzował grupy ludzi, dla których wysiłek fizyczny będzie szczególnie ważnym i skutecznym sposobem zapobiegania chorobom układu krążenia.
Opracowała: Katarzyna Sowa-LewandowskaLiteratura:1. Astrand PO et al.: Intraarterial blood pressure during exercise with different muscle groups. J Appl Physiol 1965; 20: 253-256.
2. Biochemistry of exercise-induced metabolic acidosis, Robert A. Robergs,1 Farzenah Ghiasvand and Daryl Parker Science Program, California State University-Sacramento, Sacramento, California
3. Eisenach JH et al.: The Arg16/Gly beta2-adrenergic receptor polymorphism is associated with altered cardiovascular responses to isometric exercise. Physiol Genomics. 2004;16(3):323-8.4.
4. Enrico Arcellli i Renato Canova – Marathon Traning A Scientific Approach, 2003.
5.Górski J., Fizjologiczne podstawy wysiłku fizycznego, Wydawnictwo Lekarskie,
Warszawa 2006, wyd.2
6. Guy Thibault, François Pérnott, It is not lactic acid's fault, News Studies in Athetics, kwiecień 2006.
7. Kałka D, Sobieszczańska M, Marciniak W: Aktywność fizyczna jako element prewencji chorób sercowo-naczyniowych u osób w podeszłym wieku. Pol Merkur Lekarski. 2007; 22(127):48-53.
8..Oh SD: The distribution of I/D polymorphism in the ACE gene among Korean male elite athletes. J Sports Med Phys Fitness. 2007 47(2):250-4
9. Papadimitriou ID et al.: The ACTN3 Gene in Elite Greek Track and Field Athletes. Int J Sports Med. 2007.
10. Podstawy Fizjologi wysiłku fizycznego z zarysem fizjologii człowieka pod redakcją Artura Jaskólskiego - Wrocław 2002
11. Williams AG et al.: Bradykinin receptor gene variant and human physical performance.J Appl Physiol. 2004; 96(3):938-42.
12. Wlimore JH, Costill DL: Cardiovascular control during exercise. Physiology of sport and exercise. Human Kinetics 1994; 162-188.










Recenzje