
|

Zamknij X
|



 Peroksydaza jest enzymem rozpowszechnionym w roślinach, bakteriach i tkankach zwierzęcych, reprezentującym dużą rodzinę enzymów zawierających w swym składzie hem, dzięki czemu enzym ten stosowany był w wielu badaniach analitycznych (m.in. w diagnostyce cukru i cholesterolu we krwi, testach immunologicznych - ELISA, konstrukcji biosensorów czy w przetwórstwie i przechowywaniu żwyności). Peroksydazy biorą udział w procesach metabolicznych takich jak: biogeneza etylenu, rozwój komórki i integralności błony, a także uczestniczą w mechanizmach obronnych wobec patogenów i różnych stresów abiotycznych. Ponadto, enzymy te moga brać udział w wielu reakcjach utleniania i bidegradacji, związanymi ze zmianą smaku, koloru, konsystencji oraz wartości odżywczych żywności. Kontrola aktywności peroksydazy ma duże znaczenie w procesie przetwarzania owoców, warzyw i ich produktów [1].
Peroksydaza jest enzymem rozpowszechnionym w roślinach, bakteriach i tkankach zwierzęcych, reprezentującym dużą rodzinę enzymów zawierających w swym składzie hem, dzięki czemu enzym ten stosowany był w wielu badaniach analitycznych (m.in. w diagnostyce cukru i cholesterolu we krwi, testach immunologicznych - ELISA, konstrukcji biosensorów czy w przetwórstwie i przechowywaniu żwyności). Peroksydazy biorą udział w procesach metabolicznych takich jak: biogeneza etylenu, rozwój komórki i integralności błony, a także uczestniczą w mechanizmach obronnych wobec patogenów i różnych stresów abiotycznych. Ponadto, enzymy te moga brać udział w wielu reakcjach utleniania i bidegradacji, związanymi ze zmianą smaku, koloru, konsystencji oraz wartości odżywczych żywności. Kontrola aktywności peroksydazy ma duże znaczenie w procesie przetwarzania owoców, warzyw i ich produktów [1].
Otrzymywanie peroksydazy z dzikiej rośliny Ricinus communis L. (Rącznik pospolity- należący do rodziny wilczomleczowatych) [1]
Liście rośliny (25 g) roztwartow moździerzu z dodatkiem 100 mM octanu sodu buforowanego (pH=6.0), z dodatkiem odczynnika Polyclar AT (PVPP) w ilości 1.0 g/ 10 g tkanki. Homogenat pzresączony przez 4 warstwy gazy odwirowano przy 16000 x g, przez 20 minut w temperaturze 4°C. Otrzymany czysty supernatant używano następnie do oznaczania aktywności peroksydazy metodą Neves i Lourenco (1998). Wszystkie procedury należy przeprowadzać w temperaturze 4°C. Surowy enzym wytrącono z próbki z wykorzystaniem lodowatego absolutnego etanolu, po czym próbkę odwirowano przy 15000 obrotów na minutę (15 minut). Otrzymaną peletkę zebrano, po czym rozpuszczono w 10 ml wody dejonizowanej [1].
Test enzymatyczny [1]
Aktywność peroksydazy określano przez pomiar zmiany absorbancji przy λ= 470 nm , z powodu utleniania o-dianizydyny ( 250 mM ) w obecności nadtlenku wodoru ( 500 nm ) w 1,0 ml mieszaniny reakcyjnej . Jedna jednostka aktywności enzymatycznej jest definiowana jako ilość enzymu wytwarzającego się 0,001 zmianę absorbancji na minutę w standardowych warunkach testowych [1].
Badanie optymalnego pH
Aktywność enzymu w funkcji pH ustalano za pomocą octanu sodu (pH 4,0 - 5,0 ) ,fosforanu sodu (pH 6,0-7,0 ) i buforu Tris-HCl (o pH 8.0-9.0 ) . Aktywność peroksydazy badano w standardowych warunkach[1]. Przeprowadzone badania pokazały, że enzym peroksydazy wykazuje maksymalną aktywność przy pH= 5.0, która zmniejsza się wraz ze wzrostem pH. Podobne wyniki zaobserwowano także w warunkach kwaśnego pH ok. 7.
Wpływ temperatury i stabilność termiczna
Wpływ temperatury na peroksydazy mierzono w zakresie temperatur od 30°C -80°C. Enzym inkubowano przez 10 minut w różnych temperaturach, po czym próbkę mierzono w odpowiednich odstępach czasu [1]. Peroksydazy izolowane z Ricinus communis L. Wykazały maksymalną aktywnośc w temperaturze 60°C, po czym aktywność zaczęła maleć po wzroście temperatury , a dalej zaobserwowano ostry spadek aktywności w temperaturze 80°C [1].
Oznaczanie aktywności peroksydazy gwajakolowej (GPOX) [2].
W metodzie tej określa się zawartość tetragwajakolu . Związek ten powstaje w wyniku reakcji utleniania gwajakolu. Możliwe jest także określanie w mieszaninie inkubacyjnej zawartości purpulogaliny - tj. związku powstającego w trakcie utleniania pirogalolu.
Jako materiał do przeprowadzenia oznaczenia można wykorzystać liście fasoli, pomidora lub igły sosny.
Wykonanie:
a) 1g wybranego materiału roślinnego należy zhomogenizować z dodatkiem 1-3 ml 0,05 M buforu octanowego o pH=5.6. Następnie, próbkę uzupełnić powyższym buforem do objętości 10 ml - całość odwirować. Po wirowaniu do powstałego osadu dodać kolejne 5 ml buforu octanowego, dokładnie wymieszać i ponownie odwirować. Po wirowaniu należy połączyć supernatanty.
b)Następnie, do próbki dodać 0,9 ml 0,05 M buforu octanowego (pH=5.6), a także 0,5 ml 0,02M roztworu gwajakolu (zamiast gwajakolu można zastosowac także 0,5 ml pirogalolu o takim samym stężeniu molowym), 100 µl ekstraktu enzymatycznego oraz 0,5 ml roztworu nadtlenku wodoru (H2O2).
c) Tak przygotowaną próbkę dokładnie wymieszać, po czym inkubować w 30°C, a po upływie 4 minut oznaczyć aktywność prbki przy λ= 470 nm, wykorzystując w tym celu spektrofotometr z komorą termostatową. W przypadku zastosowania roztworu pirogalolu aktywność oznaczyć przy 430 nm.
d)Równolegle do oznaczenia wykonuje się próbę kontrolną, gdzie zamiast nadtlenku wodoru dodaje się wodę. Różnica między absorbancją próbki z nadtlenkeim wodoru i bez nadtlenku stanowi miarę aktywności enzymu. Milimolowy współczynnik absorbancji tetragwajakolu wynosi 26,6mM -1cm -1, z kolei dla purpulogaliny jest on równy 2,47mM -1cm -1[2].
Przeprowadzone badania sugerują, że peroksydazy należące do ogromnej rodziny enzymów roślinnych, odgrywają rolę w procesie lignifikacji (drewnieniu) (Whetten i wsp., 1998), a także w wielu innych procesach, w tym m.in. ochronie roślin przed patogeniami czy tolerancja na sól. Określenie poszczególnych funkcji enzymów należących do peroksydaz jest niezwykle trudne ze względu na niską specyficznośc substratową w warunkach in vitro i w obecności wielu izoenzymów. Sam termin „peroksydaza” odnosi się do enzymów katalizujących reakcję oksydoredukcji (proces utleniania i redukcji , w którym następuje wymiana elektronów), zachodzący pomiędzy nadtlenkiem wodoru (H2O2), a reduktantem [3]. Jak już wspomniano, peroksydazy biorą udział w wielu procesach fizjologicznych w roślinach, obejmując odpowiedzi na stresy biotyczne i abiotyczne oraz biosyntezę ligniny. Lignina jest polimerem odpowiedzialnym za powstawanie silniejszych i stywniejszych ścian komórkowych w roślinach. Dzięki temu są one odporniejsze (usztywnione) chroniąc tym samym rośliny przed uszkodzeniami. Peroksydazy mogą być żródłem nadtlenku wodoru, ale także mają zdolność usuwania go z komórek [4].
Izolacja peroksydazy z brzoskwini (wg Neves, 2002).
Jako materiał do izolacji enzymu zastosowano brzoskwinie (Prunus persica L.), które uprawiano w warunkach standardowych. Owoce zebrano w dojrzałym stadium, umyto, obrano i pokrojono n a kawałki, po czym przechowywano w temperaturze -20 °C ( następnie używano ich do ekstrakcji enzymów) [5].
Ekstrakcja enzymu z owoców brzoskwini (wg Neves, 2002):
1) 25 g tkanki homogenizowano w 0,1M fosforanie potasu o pH=6.5 zawierającym 1mM L-cysteinę z dodatkiem Polyclar aT (0,1 g/ 10 g tkanki). Następnie, zawiesinę przesączono i zwirowano przy 25000xg przez 30 minut. Otrzymany po wirowaniu supernatant zebrano (surowy ekstrakt) i używano do oznaczania peroksydazy (POD) i PPO (peroksydaza polifenolowa) [5].
2)Frakcję osadu zawieszono w 40 ml fosforanu potasu (powyższy bufor) z dodatkiem 1M NaCl. Próbkę mieszano mechanicznie przez 12 h (przez noc), po czym odwirowano jak wyżej. Supernatand użyto jako źródło jonowego wiązania peroksydazy. W niektórych przeprowadzonych doświadczeniach NaCl zastępowano CaCl2. Całą procedurę przeprowadzono w temperaturze 4°C [5].
Test enzymatyczny [5]
Aktywność enzymu określano przez zmianę absorbancji przy λ=460 nm ze względu na utlenianie o-dianizydyny w obecności nadtlenku wodoru i enzymu. Mieszanina reakcyjna do przeprowadzenia pomiaru składała się z 0,2ml 15mM o-dianizydyny ; 0,2 ml 10mM nadtlenku wodoru; 0,1M fosforanu potasu oraz buforu cytrynianowego (o pH=5.0) w całkowitej objętości równej 3 ml.
Oczyszczanie enzymu:
a) Supernatant zawierający związaną POD po ekstrakcji dializowano wobec stałego PEG-20000 do koncentratu białka. Po zatężeniu, ekstrakt naniesiono na kolumnę z DEAE-celulozą (o wymiarach 1,5 cm x 20 cm), uprzednio zrównoważoną i przemytą za pomocą 5 mM buforu fosforanowego o pH= 7,0 [5].
b) Białko eluuje się liniowym gradientem chlorku potasu (0-0,4 M), w 5 mM buforze fosforanu potasu, o pH 7,0 w postaci 5-ml frakcji. Następnie, aktywne frakcje połączono i wprowadzono do kolumny Sephadex G-100 (o wymiarach 2 x 50 cm) zrównoważonej i przemytej tym samym buforem (jak wyżej). Eluowane frakcje zawierające aktywną peroksydazę połączono i wykorzystywano do dalszych badań [5].
Oznaczanie stężenia białka: zazwyczaj stosuje się metodę Lowry’ego, gdzie wykorzystuje się albuminę surowicy bydlęcej jako standard (pomiar przy 280 nm) [5].
Termiczna inaktywacja
Peroksydaza izolowana z owoców brzoskwini wykazuje optimum działania w temperaturze 60°C, zaś po wystawieniu na działanie temperatury równej 65°C wykazuje 45% pozostałej aktywności. W temperaturze 70°C enzym traci około 95% początkowej aktywności (po 3-minutowej inkubacji), oraz po 1-minucie w temperaturze 75°C [5].
Jak już wspomniano peroksydazy roślinne pełnią wiele fizjologicznych funkcji, ponadto znane są także zastosowania tych enzymów w praktyce (m.in. do konstruowania biosensorów stosowanych do oznaczania markerów nowotworowych, rutyny itd). Wykorzystywane są również jako elementy do zestó służących do oznaczeń immunologicznych, syntez organicznych, a także biotransformacji skłądników organicznych, gdzie najczęściej wykorzystuje się peroksydazę izolowaną z korzeni chrzanu. Aktualnie, prowadzone są poszukiwania alternatywnego źródła peroksydazy, gdzie na uwagę zasługuje peroksydaza izolowana z główek kapusty wiosennej. Może być ona potencjalny alternatywnym źrółem tego enzyu, mającym coraz szersze zastosowanie w różnych dziedzinach. Prowadzone aktualnie badania skupiają się na określeniu właściwości biochemicznych i optycznych peroksydazy izolowanej z kapusty.
Izolacja peroksydaz z kapusty (Brassica oloeracea var. capitata)
Homogenat otrzymany z główek kapusty poddaje się dwu -etapowej precypitacji z wykorzystaniem 40-75% roztworu (NH4)2SO4. Uzyskany precypitat (który zawiera całość aktywności peroksydazy z homogenatu) poddano odsoleniu przez dializę do 5mM buforu fosforanowego o pH=7.0, a następnie do wody. Na koniec próbkę zliofilizowano.
Otrzymany liofilizat nanoszono na kolumnę (o wymiarach 2,5 x 10 cm), wypełnioną DEAE-Sepharosą CL-6B . Niezwiązane na kolumnie białka wymywano wykorzystując w tym celu liniowy gradient 5-250 mM NaCl w buforze startowym. Frakcje, które wykazywały aktywność peroksydazy zebrano, a następnie odsolono i zliofilizowano.
Oznaczanie aktywności enzymu izolowanego z kapusty:
Aktywnośc peroksydazy oznaczano wobec gwajakolu. Jako jednostkę aktywności zdefiniowano taką aktywność, która w ciągu jednej minuty katalizuje utlenienie 1 µmola gwajakolu w podanych warunkach reakcji.
Do oznaczenia specyficzności substratowej wykorzystano 0,4 mM gwajakol, 0,17 mM o-dianizydynę oraz 0, 006 mM roztwór ABTS (2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid ) i 5,5 mM pirogallol [6].
Zmiany aktywności peroksydazy w odpowiedzi na zasolenie podłoża NaCl [11]
Na zmianę aktywności enzymów roślinnych mają wpływ różne czynnik, w tym np. stopień zasolenia podłoża. Biczak i wsp. (2002), przeprowadzili badania wazonowe, podczas których określano wpływ zróżnicowanego zasolenia podłoża na aktywność dwóch enzymó roślinnych tj. katalazy oraz peroksydazy. Badanie przeprowadzono na liściach selera naciowego (Apium graveolens L. var. dulce) oraz fasoli szparagowej (Phaseolus vulgaris L.) [11].
Przebieg doświadczenia:
Sadzonki obydwu roślin podlano jednorazowo roztworami wodnymi, które zawierały wzrastające stężenie NaCl tj. 30 mM NaCl, 60 mM, 120 mM i 180 mM NaCl. W uzyskanych wynikach zaobserwowano, że niezależnie od stopnia zasolenia podłoża obserwowano spadek aktywności enzymu katalazy oraz jednoczesny wzrost aktywności peroksydazy (wraz z wiekiem rośliny). Zarówno w przypadku fasoli, jak i selera naciowego, spadek aktywności katalazy był tym większy, im bardziej zasolona była ich gleba. Peroksydaza w liściach roślin, które uprawiane były na podłożu zasolonym,wykazywała wyższą aktywność w stosunku do liści roślin kontrolnych. Stopień zaobserwowanych zmian dla selera i fasoli szparagowej zależny był nie tylko od gatunku warzywa, lecz również od poziomu zasolenia podłoża (stężenie mM NaCl), oraz terminu analizy, jednakże pomijając te różnice największy wpływ na aktywność enzymów miało analizowane w trakcie doświadczenia zasolenie [11].
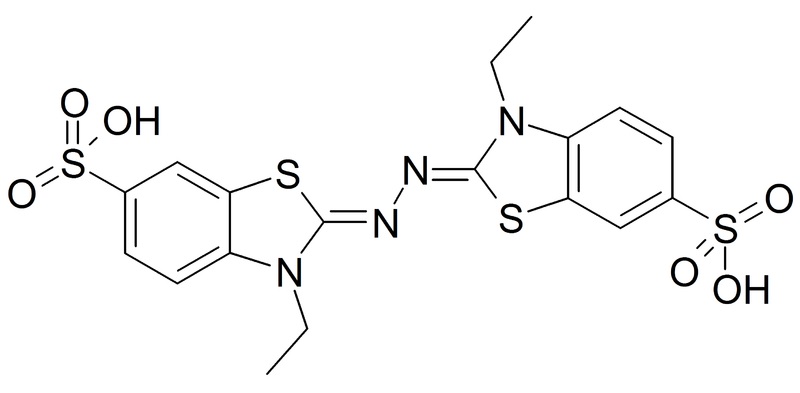
Zdjęcie: wzór ABTS, http://en.wikipedia.org/wiki/File:ABTS.png
ABTS – to kwas 2,2’-azyno-bis (3-etylenobenzotiazolinowy). Należy on do syntetycznych kationorodników, które są rozpuszczalne w wodnych i organicznych roztworach. Rodnik ten pozwala na pomiar przeciwutleniaczy zarówno hydrofilowych, jak i hydrofobowych, ponadto jest wrażliwy na światło, a po sporządzeniu musi być przetrzymywany w ciemności. W przeciwieństwie do DPPH rodnik ten nie jest dostępny w formie aktywnej- w handlu występuje pod postacią nieaktywnej soli dwuamonowej. Związek ten generuje się albo poprzez reakcję chemiczną (np. z dwutlenkiem manganu, nadsiarczanem potasu, ABAP) lub enzymatyczną (z peroksydazą, mioglobiną, hemoglobiną) [7].
Przygotowanie roztworu podstawowego ABTS [7]:
Przygotowany roztwór podstawowy przyjmuje charakterystyczny ciemno-zielony kolor [7].
Rośliny wytwarzają szereg mechanizmów obronnych (w tym produkują enzymy np. peroksydazę czy katalazę) co jest następstwem zadziałania czynnika stresowego. Odpowiedzią roślin na jego pojawienie się, jest zachowanie równowagi pomiędzy wytwarzaniem reaktywnych form tlenu (RFT) , a ich usuwaniem, z wykorzystaniem specjalnych systemów. To właśnie reaktywne formy tlenu powodują, że rośliny wykształcają mechanizm obronny w postaci produkcji np. odpowiednich enzymów [9]. Równowaga między poziomem reaktywnych form tlenu, a aktywnością komórkowego systemu antyoksydacyjnego określana jest mianem bariery antyoksydacyjnej. W systemie tym uczestniczą, oprócz peroksydazy także inne dobrze znane enzymy, w tym: dysmutaza ponadtlenkowa czy katalaza [10].
Oznaczanie zdolności antyoksydacyjnej
Przeciwrodnikowe (antyoksydacyjne) właściwości różnych roztworów wodnych można oznaczyć dzięki badaniu ich zdolności do dezaktywacji kationorodników ABTS•+. Ogólnie, zasada tej metody polega na bezpośrednim generowaniu ABTS•+ w wyniku utleniania ABTS przez działanie nadsiarczanem potasu. Kationorodnik ABTS•+ w przeciwieństwie do ABTS- u charakteryzuję się bardzo intensywnym zabarwieniem. Dodanie odpowiedniego przeciwutleniacza do próbki, prowadzi do redukcji kationorodnika ABTS•+ do ABTS, a także spadek intensywności zabarwienia roztworu rodników. Stopień redukcji ABTS•+ określany jest za pomocą spektrofotometru - pomiar przy długości fali równej λ=734 nm [8].
Przygotowanie roztworu roboczego ABTS (ABTS0.7)
Roztwór podstawowy ABTS należy rozcieńczyć za pomocą roztworu PBS tak, by absorbancja ABTS przy λ= 734 nm wynosiła A= 0,7 ± 0,02. Najlepiej by uzyskany wynik był bliższy wartości 0,72 [7].
Peroksydaza jest produkowana
także przez drożdże Streptomyces sp. W
optymalnych warunkach hodowlanych tj. w temperaturze 37°C, pH=8.8 w obecności 0,1mmol l-1
alkoholu weratralowego (3,4-dimetoksybenzyl), Musengi A. I wsp.(2013)
zaobserwowali najwyższą aktywność peroksydazy. Obserwacje te prowadzono w 10 ml
objętości hodowli, zaś skalowanie do 100 ml i 400 ml objetości hodowli
powodowało zmniejszenie produkcji peroksydazy [12]. Przeprowadzone badania wykazały,że alkohol weratrylowy jest
bardzo dobrym induktorem produkcji peroksydazy przez Streptomyces sp. [12].
Autor: Lidia Koperwas
Literatura:
[1]. Isolation and characterization of peroxidase from the leaves of Ricinus communis., 2008. http://www.thefreelibrary.co/Isolation+and+characterization+of+peroxidase+from+the+leaves+of...-a0215925289
[2]. Kłyszejko-Stefanowicz L, 2003. Ćwiczenia z biochemii. Wydawnictwo Naukowe PWN, 2003, s.543
[3]. Oxford Journals, 2013. A Large Family of Class III Plant Peroxidases. Oxford Journals , Life Sciences,Plant and Cell Physiology , Volume 42, Issue 5 , Pp. 462-468. http://pcp.oxfordjournals.org/content/42/5/462.full
[4]. Vicuna, Deborah (Thesis), "The role of peroxidases in the development of plants and their responses to abiotic stresses." (2005). Doctoral. Paper 15. http://arrow.dit.ie/sciendoc/15
[5]. Neves V.,A., 2002. Ionically Bound Peroxidase from Peach Fruit. Brazilian Archives of Biology and Technology,an International Journal, Vol. 45, N. 1 : pp. 7 ,16, March, 2002, ISSN 1516-8913 Printed in Brazil
[6]. Belcarz A., Ginalska G., Kowalewska B., Kulesza P., 2007. Peroksydaza z kapusty i jej zastosowanie w multifinkcjonalnych biosensorach. XV KbiIB 2007. http://www.chem.uw.edu.pl/pbz18/group_papers/belcarz1.pdf
[7].http://matrix.ur.krakow.pl/~aduda-chodak/dydaktyka/BIOAKTYWNE/cwiczenie%20I%20praktyka.pdf
[8]. http://zbm.umed.pl/doki/Rodniki/cwiczenia/cw2.pdf
[9]. Furmanek T., 2008. Odpowiedź antyoksydacyjna roślin pomidora na infekcję Botrytis Cinerea. Słup s k i e P r a c e B i o l o g i c z n e 5 , 2008. http://apsl.edu.pl/spb/pliki/nr5/furmanek.pdf
[10]. Puzanowska-Tarasiewicz H., Kuźmicka L., Tarasiewicz M., 2009. Obrona organizmu przed reaktywnymi formami tlenu. Wiadomości lekarskie, 2009,LXII,4; 248-256. http://wiadomoscilekarskie.pl/pl/articles/item/18000/obrona_organizmu_przed_reaktywnymi_formami_tlenu
[11]. Biczak R., Herman B., Rychter P., 2002. Wpływ zasolenia NaCl na aktywność katalizy i peroksydazy w liściach selera naciowego i fasoli szparagowej. Chemia i Inżynieria Ekologiczna 2002 | Vol. 9, nr 2-3 | 205-211
[12]. Musengi A, Khan N, Le Roes-Hill M, Pletschke BI, Burton SG, 2013. Increasing the Scale of Peroxidase Production by Streptomyces sp. strain BSII#1. J Appl Microbiol. 2013 Oct 31. doi: 10.1111/jam.12380. http://www.ncbi.nlm.nih.gov/pubmed/24176016
25 maja 2018 roku zacznie obowiązywać Rozporządzenie Parlamentu Europejskiego i Rady (UE) 2016/679 z dnia 27 kwietnia 2016 r (RODO). Potrzebujemy Twojej zgody na przetwarzanie Twoich danych osobowych przechowywanych w plikach cookies. Poniżej znajdziesz pełny zakres informacji na ten temat.
Zgadzam się na przechowywanie na urządzeniu, z którego korzystam tzw. plików cookies oraz na przetwarzanie moich danych osobowych pozostawianych w czasie korzystania przeze mnie ze strony internetowej Laboratoria.net w celach marketingowych, w tym na profilowanie i w celach analitycznych.
Administratorami Twoich danych będziemy my: Portal Laboratoria.net z siedzibą w Krakowie (Grupa INTS ul. Czerwone Maki 55/25 30-392 Kraków).
Chodzi o dane osobowe, które są zbierane w ramach korzystania przez Ciebie z naszych usług w tym zapisywanych w plikach cookies.
Przetwarzamy te dane w celach opisanych w polityce prywatności, między innymi aby:
dopasować treści stron i ich tematykę, w tym tematykę ukazujących się tam materiałów do Twoich zainteresowań,
dokonywać pomiarów, które pozwalają nam udoskonalać nasze usługi i sprawić, że będą maksymalnie odpowiadać Twoim potrzebom,
pokazywać Ci reklamy dopasowane do Twoich potrzeb i zainteresowań.
Zgodnie z obowiązującym prawem Twoje dane możemy przekazywać podmiotom przetwarzającym je na nasze zlecenie, np. agencjom marketingowym, podwykonawcom naszych usług oraz podmiotom uprawnionym do uzyskania danych na podstawie obowiązującego prawa np. sądom lub organom ścigania – oczywiście tylko gdy wystąpią z żądaniem w oparciu o stosowną podstawę prawną.
Masz między innymi prawo do żądania dostępu do danych, sprostowania, usunięcia lub ograniczenia ich przetwarzania. Możesz także wycofać zgodę na przetwarzanie danych osobowych, zgłosić sprzeciw oraz skorzystać z innych praw.
Każde przetwarzanie Twoich danych musi być oparte na właściwej, zgodnej z obowiązującymi przepisami, podstawie prawnej. Podstawą prawną przetwarzania Twoich danych w celu świadczenia usług, w tym dopasowywania ich do Twoich zainteresowań, analizowania ich i udoskonalania oraz zapewniania ich bezpieczeństwa jest niezbędność do wykonania umów o ich świadczenie (tymi umowami są zazwyczaj regulaminy lub podobne dokumenty dostępne w usługach, z których korzystasz). Taką podstawą prawną dla pomiarów statystycznych i marketingu własnego administratorów jest tzw. uzasadniony interes administratora. Przetwarzanie Twoich danych w celach marketingowych podmiotów trzecich będzie odbywać się na podstawie Twojej dobrowolnej zgody.
Dlatego też proszę zaznacz przycisk "zgadzam się" jeżeli zgadzasz się na przetwarzanie Twoich danych osobowych zbieranych w ramach korzystania przez ze mnie z portalu *Laboratoria.net, udostępnianych zarówno w wersji "desktop", jak i "mobile", w tym także zbieranych w tzw. plikach cookies. Wyrażenie zgody jest dobrowolne i możesz ją w dowolnym momencie wycofać.
Więcej w naszej POLITYCE PRYWATNOŚCI




{kind=link}
Recenzje